06.03.2014
Эпителиально-мезенхимальный переход (Epithelial-mesenchymal transition, EMT) — сложный процесс изменения эпителиальными клетками эпителиального фенотипа на мезенхимальный, происходящий в эмбриональном развиитии, заживлении ран, а также при патологических процессах — например, при фиброзе, а также при опухолевой прогрессии.
Существует также и обратный процесс — мезенхимально-эпителиальный переход (Mesenchymal-epithelial transition).
Среди механизмов EMT можно выделить несколько ключевых моментов:
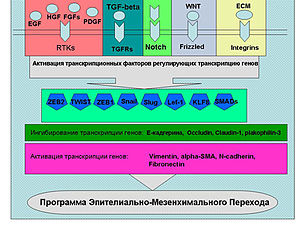
Растворимые факторы роста (на схеме), цитокины, молекулы внеклеточного матрикса активируют сигнальные пути ведущие к реализации программы ЭМП. Эти пути активируют ряд транскрипционных факторов (Snail, Twist, Slug, ZEB1, ZEB2, Lef-1 и др.), которые связываются с промоторами генов ответственных за ЭМП.
Промоторы генов, кодирующих белки плотных контактов (Tight Junction)(TJ)) - E-cadherin, occludin, claudin-1, транскрипционно ингибируются этими транскрипционными факторами, а соответственно промоторы генов компонентов цитоскелета, например Vimentin, а также генов белков внеклеточного матрикса Fibronectin, в свою очередь, наоборот активируются.
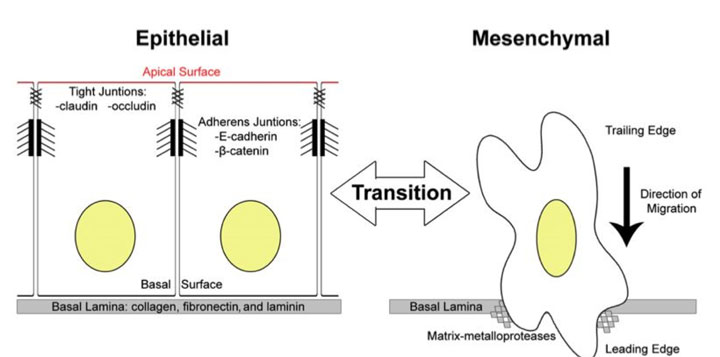
Одним из основных процессов, происходящих во время регенерации тканей является превращение эпителиальных клеток в мезенхимальные и наоборот, из мезенхимальных клеток в эпителиальные.
На рисунке выше показана разница между этими двумя основными фенотипами клеток. Важно, что нет никаких других многоклеточных тканей. Эпителиальные клетки плотно связаны друг с другом и с внеклеточным матриксом. Внеклеточный матрикс является базальной пластинкой, которая служит своего рода «колыбелью» для эпителиальных клеток. Мезенхимальные клетки расположены в 3D внеклеточной матрицы. Они биполярны, а это значит, что у них есть другое расположение цитоскелета и распределение органелл внутри них.
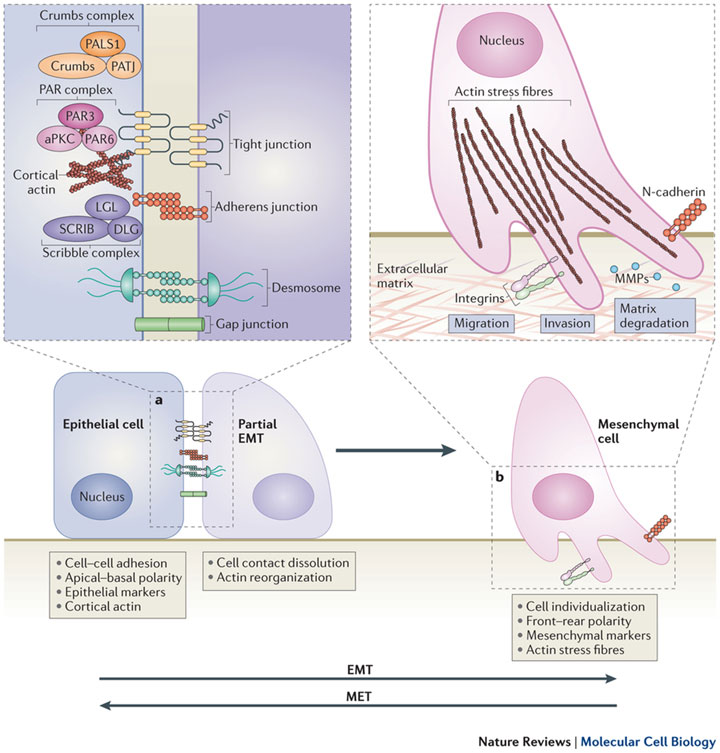
а. Первый этап эпителиально - мезенхимального перехода (epithelial-mesenchymal transition - ЕМT) – разрушение эпителиальных межклеточных контактов, плотных соединений, слипающихся между собой, десмосом и щелевых контактов , а также, нарушение клеточной полярности через нарушение частиц , разделение неисправных так называемых PAR- и Scribble (SCRIB ) -полярные комплексы. Экспрессия эпителиальных генов подавляется, одновременно с активацией экспрессии мезенхимальных генов.
б. Далее эпителиальная структура актина реорганизуется и клетки приобретают подвижность и инвазивный потенциала путем формирования ламеллиподий , филоподий и инвадопий , а также, с помощью экспрессии матричной металлопротеиназы (matrix metalloproteinases - MMPs), что, в дальнейшем, может привести к снижению внеклеточных матричных белков (extracellular matrix (ECM) proteins. Процесс мезенхимально -эпителиального перехода (МЕТ) позволяет клеткам , которые подверглись EMT вернуться в эпителий.
Утрата характерных черт эпителия (структуры ткани, клеточных взаимодействий, контроля специфическими факторами роста, приобретение подвижности и морфологии фибробластов) – это так называемое EMT, эпителиально-мезенхимальное превращение. ЕМТ свойственно нормальному эпителию в процессе развития, особенно раннего, например при гаструляции, когда эпителий приобретает подвижность и активно внедряется в подлежащие слои. ЕМТ имеет место при временных повреждениях ткани, при этом эпителиальные клетки теряют полярность, прекращают синтез кадхеринов, образуют виментин и фибронектин и одновременно с этим приобретают подвижность. Они прекращают синтез клеточных ядерных трансфакторов и образование антигенов, характерных для эпителиальных тканей. Эпителиальные клетки становятся типичными фибробластами. ЕМТ, по-видимому, лежит в основе инвазии и метастазирования: клетки эпителиальной опухоли становятся подвижными и приобретают способность расселяться по разным территориям организма. При этом очень существенно, что клетки претерпевают физиологическое, а не генетическое превращение, так как ЕМТ обратимо.
Метастазы, возникшие на основе ЕМТ, могут приобретать морфологию исходной опухоли, а эпителий в краевых районах раны может приобретать фибробластные свойства. Индукция ЕМТ имеет место при взаимодействии опухолей, экспрессирующих онкоген Ras и TGFр. Но так или иначе ЕМТ выглядит как заключительный этап прогрессии эпителиальной опухоли, когда опухоль теряет эпителиальные признаки (полярность клеток, специфические клеточные контакты, характерную морфологию и тканеспецифическую антигенную структуру) и одновременно приобретает черты фибробластов (экспрессию виментина, подвижность, независимость от территории роста).
Можно думать, что понимание этого процесса и факторов, в нем участвующих, создадут основу для рациональной терапии инвазии и метастазирования – главных свойств злокачественности. При этом непонятно, что будет дальше. Ведь прогрессия должна быть бесконечна, а EMT как бы завершает ее.
Источник: Nature Reviews Molecular Cell Biology 15,178–196, (2014)