Копнин Б.П.
ФГБУ «НМИЦ онкологии им. Н.Н. Блохина» Минздрава России, Москва
1.1. Характерные признаки опухолевой клетки
Злокачественные новообразования возникают в результате неограниченной пролиферации клеточного клона, выходящего за пределы собственной ткани и способного к росту на территориях других тканей. При этом в силу высокой генетической изменчивости и селекции, происходящей под давлением со стороны организма, в популяции клеток такого клона постоянно возникают и отбираются все более и более автономные и агрессивные субклоны, что описывается термином опухолевая прогрессия. В результате довольно длительной эволюции неопластического клона формируется опухоль, способная убить организм. В последнее десятилетие был достигнут значительный прогресс как в идентификации генов, нарушения функции которых ведут к развитию новообразований, так и в выяснении роли белковых продуктов таких генов в физиологии клетки. Все это позволило выделить ряд важнейших свойств, приобретение которых предопределяет способность клетки образовывать злокачественную опухоль
(Рис.1).
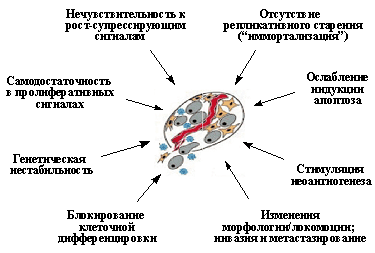
Во-первых, это пониженная потребность во внешних сигналах для инициации и поддержания клеточной пролиферации - так называемая самодостаточность в пролиферативных сигналах. Данное положение может быть проиллюстрировано двумя примерами (Рис. 2). При культивировании in vitro большинство типов нормальных клеток размножается лишь при условии, если питательная среда содержит 10-20% сыворотки, т.е. при довольно значительном содержании в ней различных ростовых факторов (Рис. 2А). Связывание ростовых факторов со своими рецепторами инициирует передачу сигналов внутри клетки, приводящую к репликации ДНК и делению клетки. Оказалось, что многие типы опухолевых клеток способны размножаться в среде с 1% и даже 0,1% сыворотки, т.е. при содержании ростовых факторов в десятки и сотни раз меньшем, чем необходимо для стимуляции размножения нормальных клеток. Такая пониженная потребность в растворимых ростовых факторах достигается изменениями в системах внутриклеточной сигнализации, которые либо вызывают секрецию необходимых факторов роста самими трансформированными клетками, либо резко увеличивают количество рецепторов для необходимых факторов роста, либо запускают в отсутствии ростового фактора каскад событий, аналогичный тому, который в норме инициируется связыванием ростового фактора со своим рецептором.
Другим примером пониженной потребности неопластических клеток во внешних пролиферативных сигналах является их так называемая независимость от субстрата (anchorage-independence) - Рис. 2Б. Большинство типов нормальных клеток способны размножаться лишь при условии их прикрепления к определенному внеклеточному матриксу. Например, фибробласты начинают делиться при взаимодействии с фибронектином. В ином случае пролиферативный стимул, исходящий от растворимых ростовых факторов, не вызывает полноценного каскада передачи внутриклеточных сигналов, необходимого для стимуляции размножения клеток. Многие типы опухолевых клеток, в отличие от их нормальных предшественников, способны пролиферировать, не прикрепляясь к субстрату, например, в полужидкой среде. Эти два примера показывают, что неопластические клетки приобретают способность генерировать внутри себя пролиферативные сигналы, в норме исходящие от внешних стимулов.
Вторым важнейшим приобретенным свойством неопластических клеток является их пониженная чувствительность к рост-ингибирующим сигналам. Как известно, в организме существует множество антипролиферативных сигналов, поддерживающих определенное число клеток в каждой из тканей. Такие сигналы генерируются как секретируемыми растворимыми факторами (цитокинами), так и взаимодействиями клеток с внеклеточным матриксом и друг с другом. Классическим примером здесь является так называемое контактное торможение размножения клеток в культурах in vitro. (Рис. 2В).
Нормальные клетки, например фибробласты, размножаются до тех пор, пока не возникнет плотный монослой и не установятся межклеточные контакты. В отличие от этого, трансформированные клетки при возникновении межклеточных контактов не останавливают свою пролиферацию, а продолжают делиться, наползать друг на друга и образовывать очаги многослойного роста.
А) Пониженная зависимость от факторов роста

Б) Способность размножаться без прикрепления к внеклеточному матриксу

В) Отсутствие контактного торможения размножения
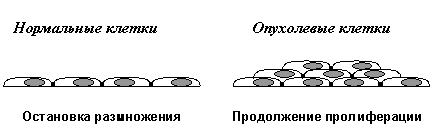
Рис.2. Характерные признаки неопластических клеток in vitro (объяснения в тексте)
Наряду с этим, опухолевые клетки, как правило, значительно менее чувствительны к действию рост-ингибирующих цитокинов, факторов специфического и неспецифического противоопухолевого иммунитета, а кроме того, не останавливают свою пролиферацию при ДНК-повреждающих воздействиях или неблагоприятных условиях - недостатке пула нуклеотидов, гипоксии и т.д.
Еще одним важнейшим свойством опухолевых клеток является отсутствие репликативного старения, или приобретение бессмертия (иммортализация). Как известно, существует механизм, ограничивающий число делений большинства типов зрелых клеток человека. Так, в культурах человеческих фибробластов in vitro после 60-80 делений (так называемое число Хейфлика) наблюдается необратимая остановка размножения клеток и их постепенная гибель. Между тем, чтобы образовать из одной клетки-родоначальницы сначала опухоль, а затем и метастазы, в условиях жесткого давления со стороны организма, когда многие опухолевые клетки погибают, может потребоваться большее число делений. И, действительно, в опухолевых клетках наблюдается нарушение работы такого "счетно-ограничительного" механизма контроля репликации.
Следующим важным свойством неопластических клеток является ослабление индукции в них апоптоза. Апоптоз представляет собой активный механизм клеточного самоубийства, поддерживающий в организме определенное число клеток и, кроме того, защищающий его от накопления аномальных клеточных вариантов. Он вызывается как физиологическими сигналами (связыванием специфических киллерных цитокинов со своими рецепторами), так и различными внутриклеточными повреждениями или неблагоприятными условиями, в частности нарушениями структуры ДНК, нехваткой ростовых факторов, гипоксией и т.д. Уход от апоптоза резко повышает жизнеспособность неопластической клетки, делает ее менее чувствительной к факторам противоопухолевого иммунитета и терапевтическим воздействиям.
К важнейшим приобретенным свойствам опухолевых клеток принадлежит и их способность стимулировать неоангиогенез, т.е. формировать новые кровеносные и лимфатические сосуды из эндотелиальных клеток предсуществующих окружающих мелких сосудов. Это необходимое условие для дальнейшего роста опухолевого узелка, достигшего в диаметре 2-4 мм. В ином случае клетки в центре опухоли, не получая кислород и питательные вещества, будут погибать.
Важнейшим свойством опухолевых клеток являются и изменения морфологии и движения клеток. В основе морфологических нарушений лежат взаимосвязанные между собой изменения цитоскелета, адгезионных взаимодействий клеток друг с другом и с внеклеточным матриксом. Вкратце, они выражаются в нарушении формирования фокальных контактов и ухудшении прикрепления клеток к внеклеточному матриксу, дезорганизации системы актиновых микрофиламентов. Это приводит к изменениям активности псевдоподий и подвижности. В целом, наблюдаемая картина напоминает изменения, возникающие в нормальных клетках при действии мотогенных цитокинов - факторов, стимулирующих миграцию клеток. Однако так называемый локомоторный фенотип в неопластических клетках, как правило, сильно утрирован, что позволяет различать по морфологии опухолевую клетку от движущейся нормальной клетки. Необходимо подчеркнуть, что именно эти нарушения, вместе с некоторыми другими свойствами, в частности способностью секретировать протеолитические энзимы, предопределяют приобретение неопластическими клетками двух свойств, лежащих в основе злокачественного роста: способность к инвазии, т.е. проникновению в окружающие здоровые ткани, и сопряженную с ней способность к метастазированию - образованию вторичных очагов опухолевого роста. Метастазирование - наиболее опасное проявление опухолевой прогрессии, являющееся основной причиной смерти онкологических больных. Чтобы дать метастаз клетка должна приобрести ряд свойств: умение проникать в глубину окружающих нормальных тканей, в том числе в кровеносные или лимфатические сосуды; способность выживать после попадания в сосуды, а затем - выходить из них и размножаться в несвойственном для данного типа клеток микроокружении, давая новый очаг опухолевого роста. Таким образом, способность к метастазированию складывается из комплекса более простых признаков, таких как приобретение локомоторного фенотипа, способности стимулировать образование новых кровеносных и лимфатических сосудов, создавая тем самым пути эвакуации опухолевых клеток из первичного очага, возникновение независимости от субстрата, подавление апоптоза и т.д. Появление каждого из этих свойств увеличивает метастатический потенциал клетки.
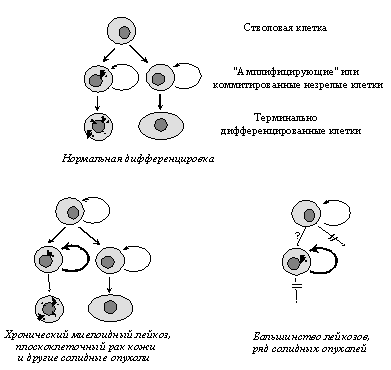
Для многих опухолевых клеток характерны и нарушения клеточной дифференцировки, т.е. образования специализированных типов клеток, синтезирующих специфические белки. Особенно ярко это проявляется в гемобластозах, новообразованиях из кроветворных тканей, при которых их клетки оказываются как бы замороженными на той или иной стадии созревания. Общепринятым является представление, согласно которому меньшая зрелость лейкозных клеток является не следствием дедифференцировки зрелых клеток, претерпевших неопластическую трансформацию, а отражает их происхождение из незрелых клеток, в которых блокированы процессы дальнейшей дифференцировки. Следует заметить, однако, что это свойство не является универсальным: во многих типах опухолей наблюдается сохранение способности к дифференцировке, причем в отличие от лейкозов созревание клеток не препятствует приобретению злокачественного фенотипа. Примерами этого могут служить плоскоклеточный ороговевающий рак кожи и высокодифференцированные аденокарциномы толстой кишки, происходящие из незрелых клеток, которые сначала несколько раз делятся, а затем дифференцируются (Рис. 3).
Происхождение из незрелых клеток не противоречит представлению о том, что опухолевые клетки в ходе прогрессии могут претерпевать определенную дедифференцировку, утрачивая в первую очередь те дифференцировочные белки, отсутствие которых придает клеткам селективные преимущества (например, рецепторы стероидных гормонов в раках молочной железы и т.д.).
И, наконец, важнейшим признаком неопластических клеток является их генетическая нестабильность. Очевидно, что канцерогенез - многоступенчатый процесс накопления мутаций и других генетических изменений, приводящих к нарушениям регуляции размножения и миграции клеток, понижению их чувствительности к различным рост-супрессирующим сигналам, ослаблению индукции в них апоптоза, блокированию дифференцировки и т.д. Вероятность возникновения в одной клетке нескольких генетических изменений, придающих совокупность вышеуказанных свойств, резко повышается при нарушениях работы систем, поддерживающих целостность генома. Поэтому мутации, ведущие к генетической нестабильности, также являются неотъемлемым этапом опухолевой прогрессии. Генетическая нестабильность неопластических клеток базируется на уменьшении точности воспроизведения генетического аппарата, нарушениях механизмов репарации ДНК и изменениях регуляции клеточного цикла в поврежденных клетках. Это, вместе с уходом от апоптоза, позволяющим генетически измененным клеткам выживать, делает популяции опухолевых клеток высоко изменчивыми, создает основу для постоянного возникновения и отбора все более и более злокачественных вариантов. Таким образом, генетическая нестабильность является двигателем неуклонной опухолевой прогрессии.